Ymp‰ristˆn aiheuttamat
muutokset voivat kest‰‰
useita sukupolvia
Paramecium caudatum
|
Tiensuu, Sampo; Dauermodifikaatio - Ymp‰ristˆn aiheuttamat muutokset voivat kest‰‰ useita sukupolvia. Seepia 5 (2002):3-7 |
|
|
Ymp‰ristˆn aiheuttamat
|
|
Paramecium caudatum |
Ennen kuin Charles Darwinin ajatus luonnonvalinnasta tuli yleisesti hyv‰ksytyksi, oli vallalla ranskalaisen Jean Baptiste Lamarckin (1744-1829) teoria lajien sopeutumisesta. Lamarckin mukaan kaikki eliˆt pystyv‰t muuttamaan itse‰‰n elinymp‰ristˆˆns‰ paremmin sopeutuviksi ja siirt‰m‰‰n uudet ominaisuudet j‰lkel‰isilleen. Teoksessaan The Origin of Species (1859, suom. Lajien synty) Darwin kumosi Lamarckin v‰itteet laajan todistusaineistonsa perusteella ja esitti vaihtoehtoisen luonnonvalintaan perustuvan teorian, joka on toiminut myˆs nykyisen evoluutioteorian perustana. Myˆhemmin Darwin alkoi kuitenkin uusien havaintojensa perusteella yh‰ enemm‰n vakuuttua siit‰, ett‰ myˆs hankittujen ominaisuuksien periytymisell‰ saattaa olla osuutensa evoluutiossa [4]. Lukuisat kokeet osoittavat, ett‰ hankitut ominaisuudet voivat todella siirty‰ tuleville sukupolville. Niiden osuudesta evoluutiossa ei kuitenkaan ole mink‰‰n laista n‰yttˆ‰. Havainnoissa ei siis varsinaisesti ole kysymys lamarckismista.
Ven‰l‰issyntyinen saksalainen Victor Jollos havaitsi t‰llaisen ilmiˆn 1910-luvun alussa tutkiessaan tohveliel‰inten (l‰hinn‰ Paramecium caudatum, P. aurelium ja P. multimicronucleatum) kyky‰ kest‰‰ korkeita l‰mpˆtiloja ja kemikaaleja, kuten arsenikkia [7]. Kun tohveliel‰imi‰ pidettiin tarpeeksi pitk‰‰n tavallista korkeammassa l‰mpˆtilassa, niiden l‰mmˆnsietokyvyn yl‰raja nousi kaksi, joskus jopa kolme astetta samaan klooniin kuuluvaan vertailuryhm‰‰n n‰hden. (Tohveliel‰inten yhteydess‰ kloonilla tarkoitetaan kaikkia niit‰ yksilˆit‰, joilla on sama perim‰. T‰llaisia ryhmi‰ syntyy suvuttoman lis‰‰ntymisen surauksena.)
|
|
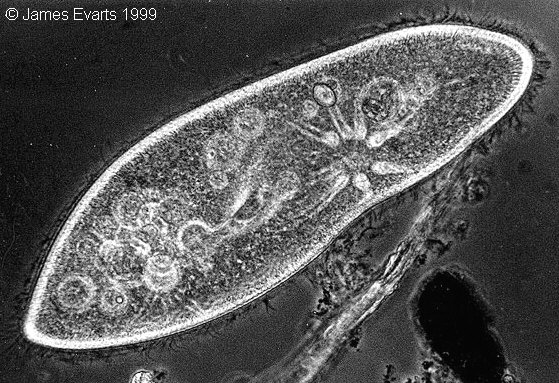
Kuva 1: Er‰s tohveliel‰in (Paramecium caudatum). Isotuma ei n‰y kuvassa, sill‰ se on juuri hajonnut er‰‰ss‰ tohveliel‰imille luontaisessa prosessissa. Kuvan kaksi sinist‰ vesikkeli‰ kehittyv‰t myˆhemmin tumiksi. |
L‰mmˆnsietokyvyn kasvaminen kesti kuitenkin useita vuosia eli tuhansia sukupolvia. T‰m‰n j‰lkeen ominaisuus s‰ilyi niin kauan kuin el‰imet lis‰‰ntyiv‰t suvuttomasti, mutta kahden tohveliel‰imen v‰linen konjugaatio eli geenien vaihto sek‰ endomiksiaksi kutsuttu tuman uudelleenj‰rjest‰ytymisprosessi yleens‰ poistivat ominaisuuden. Jos korkea l‰mpˆtila oli kuitenkin jatkunut tarpeeksi kauan, muutoksen h‰vi‰minen saattoi kest‰‰ yhden, joskus useitakin konjugaatioita. Jollos teki vastaavia kokeita myˆs arsenikilla saaden sanamkaltaisia tuloksia. Niiss‰ el‰inten kuolleisuus oli kuitenkin suuri, joten tulokset voivat ainakin osittain selitty‰ mutaatioiden ja valinnan avulla. Arsenikkiresistanssikin kuitenkin h‰visi v‰hitellen, mutta saattoi kest‰‰ jopa 600 sukupolvea. [20] Jollosin kokeet ovat varsin vakuuttavia, vaikka h‰nt‰ onkin kritisoitu siit‰, ett‰ h‰n ei kontrolloinut kunnolla ravinnon saatavuutta kokeiden aikana [20].
Jollos alkoi kutsua ilmiˆt‰ dauermodifikaatioksi (saks. Dauer=kesto). H‰n p‰‰tteli, ett‰ dauermodifikaatio siirtyy sukupolvelta toiselle solulimassa eik‰ geeneiss‰. Nyky‰‰n termi‰ k‰ytet‰‰n tarkoittamaan kaikkia ymp‰ristˆn aiheuttamia muutoksia, jotka s‰ilyv‰t sukupolvelta toiselle, vaikka muutoksen aiheuttanut ymp‰ristˆtekij‰ ei en‰‰ olisikaan l‰sn‰, mutta jotka kuitenkin lopulta h‰vi‰v‰t. Ilmiˆst‰ on myˆhemmin tehty lis‰‰ havaintoja paitsi alkuel‰imill‰ [16], myˆs monisoluisilla eliˆill‰. Niill‰ muutos kest‰‰ kuitenkin vain muutamia sukupolvia.
|
|
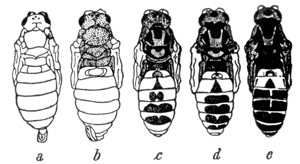
Kuva 4: Habrobracon juglandiksen v‰rimuutosasteita. Tummuminen aiheutuu alunperin korkeasta tai matalasta l‰mpˆtilasta kotelovaiheen aikana. Muutos n‰kyy kuitenkin viel‰ kahdessa seuraavassa sukupolvessa, vaikka ne kasvaisivatkin t‰ysin normaaleissa olosuhteissa. Kuva l‰hteest‰ [18]. |
Aikuisten hyˆnteisten kutikulaa v‰ritt‰v‰t pigmentit syntyv‰t yleens‰ kotelovaiheen aikana. Kun er‰‰n vainopisti‰islajin (Habrobracon juglandis) koteloita pidettiin yhden tunnin ajan korkeassa (+45 ∫C) tai matalassa (ñ13 ∫C) l‰mpˆtilassa, pigmentti‰ kehittyi tavallista enemm‰n, jonka seurauksena kuoriutuvat pisti‰iset olivat normaalia tummempia [11, 13, 18] (ks. kuva 4). V‰rimuutos n‰kyi kahdessa seuraavassa sukupolvessa vaikkakin heikentyneen‰. Kolmannessa polvessa se oli h‰vimmyt kokonaan. Muutos n‰kyi erityisesti koiraspisti‰isill‰, kun taas naarailla ero on tuskin huomattava. Merkitt‰v‰‰ on, ett‰ se kuitenkin periytyi l‰hes pelk‰st‰‰n naaraiden v‰lityksell‰. T‰m‰ n‰ytt‰isi tukevan k‰sityst‰, jonka mukaan dauermodifikaatio siirtyy solulimassa, sill‰ siittiˆn munasoluun tuoman soluliman m‰‰r‰ on v‰h‰inen.
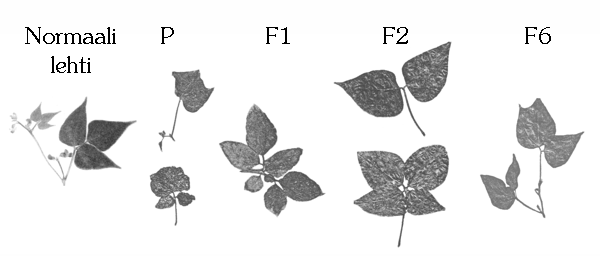
Toisen merkitt‰v‰n havainnon teki Frederick Wenzl Hofmann tarhapavulla (Phaseolus vulgaris) ñ samalla lajilla, jota tutkimalla Mendel aikoinaan kehitti periytymisoppinsa. T‰ll‰ kertaa muutos kesti kuusi sukupolvea! [6] H‰n k‰sitteli kasveja koko niiden kasvukauden ajan 0,75-prosenttisella kloraalihydraatilla, joka tunnetaan myˆs îtyrm‰ystippoinaî. Jotta geenien vaihdunnasta johtuvia fenotyypin muutoksia ei tapahtuisi, h‰n valitsi kasvit puhtaasta linjasta. Ne olivat toisin sanoen homotsygootteja kaikkien geeniens‰ suhteen. K‰sitellyiss‰ kasveissa n‰kyi lukuisia kasvuh‰iriˆit‰. Monet lehdet olivat ep‰symmetrisi‰ ja kiehkuroissa oli tavallisen kolmen sijasta kahdesta viiteen lehte‰ (ks. kuva 5). Lis‰ksi niihin ilmestyi keltaisia laikkuja.
Kasveista valittiin eniten muuntuneet ja annettiin niiden siement‰‰. Koska tarhapavut tuottavat siemenens‰ itsepˆlytyksell‰, ei perim‰n sekoittumisen vaarasta tarvinnut huolehtia. Puhtaan linjan kasvien j‰lkel‰iset ovat t‰llˆin aina perim‰lt‰‰n vanhempiensa klooneja. Siemenist‰ 200 kylvettiin ja kasvatettiin uusiksi kasveiksi. N‰ist‰ 146 oli vanhempiensa tapaan ep‰muodostuneita. N‰in jatkettaessa muuntuneitten kasvien osuus v‰heni joka sukupolvessa, kunnes seitsem‰nness‰ polvessa ei ollut en‰‰ yht‰‰n ep‰normaalia yksilˆ‰.
Hofmann osoitti risteytyskokeilla, ett‰ muutos ei ole voinut tapahtua kromosomistossa. H‰nen mukaansa îperiytyminenî ei myˆsk‰‰n ole voinut johtua kloraalihydraatista, jota olisi siirtynyt pieni‰ m‰‰ri‰ soluliman mukana, sill‰ kun kasvuh‰iriˆt johtuivat kloraalihydraatin suorasta vaikutuksesta, palautui kasvu normaaliksi heti, kun k‰sittely lopetettiin. Jos periytyminen taas olisi johtunut pelk‰st‰‰n kloraalihydraatin muuttamasta siemenkudoksesta, sen olisi pit‰nyt kest‰‰ vain yhden sukupolven, sill‰ seuraavassa sukupolvessa alkuper‰isen siemenen soluja ei en‰‰ ole j‰ljell‰. Olisi myˆs hyvin ep‰todenn‰kˆist‰, ett‰ tulokset olisivat syntyneet sattumalta, sill‰ vaikka pavuissa ilmenee luontaisesti silloin t‰llˆin t‰m‰n kaltaisia kasvuh‰iriˆit‰, niit‰ tavataan vain yhdell‰ 5000 kasvista. N‰ill‰ perusteilla Hofmann p‰‰tteli, ett‰ kyseess‰ t‰ytyy olla solulimassa tapahtunut muutos, joka aina solun jakautuessa siirtyy soluliman mukana kummallekin tyt‰rsolulle.
|
|
Kuva 5: Esimerkkej‰ tarhapavun (Phaseolus vulgaris) lehdiss‰ tapahtuneista muutoksista. Vain P-sukupolven kasveja on k‰sitelty kloraalihydraatilla, mutta myˆs kuudessa seuraavassa sukupolvessa on muuntuneita lehti‰. ñ Normaali lehti on kuvattu Helsingin yliopiston kasvimuseon kokoelmasta. Muut kuvat ovat l‰hteest‰ [6]. |
|
|
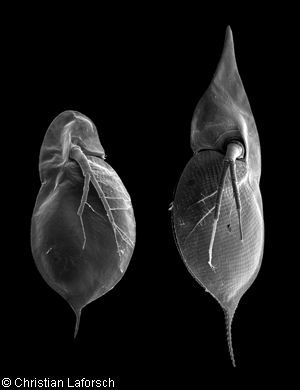
Kuva 2: Daphnia cucullata. Elektronimiksroskooppikuva, jossa n‰kyy normaali (vas.) sek‰ kairomoniymp‰ristˆss‰ syntynyt suojapanssari. Kuva 3: Daphnia longispina, joka on l‰heist‰ sukua D. cucullatalle |

|
Useita sukupolvia kest‰v‰t dauermodifikaatiot ovat kuitenkin monisoluisilla harvinaisia, vaikkakin yksisoluisilla ilmeisesti eritt‰in yleisi‰ [16, 20]. Monisoluisten eliˆiden muutoksista suurin osa periytyy ainoastaan yhden kerran. Ominaisuuden vanhemmiltaan perinyt eliˆ ei siis siirr‰ sit‰ en‰‰ omille j‰lkel‰isilleen. Yksi vaikuttavimmista esimerkeist‰ on seuraava vastik‰‰n vesikirpuilla havaittu ilmiˆ.
Jo pitk‰‰n on tiedetty, ett‰ er‰iden vesikirppujen vihollisten eritt‰m‰t kemikaalit vaikuttavat vesikirpuille kehittyv‰n suojapanssarin kokoon (ks. kuva 2). Kun vedess‰ on tiettyj‰ kairomoneiksi kutsuttuja kemikaaleja, lajista riippuen vesikirppu joko kehittyy piikikk‰‰ksi tai kasvattaa tavallista suuremman suojapanssarin. Neutraalissa ja kairomoniym‰ristˆss‰ kasvaneet vesikirput ovat kesken‰‰n niin erilaisia, ett‰ niit‰ on joskus luultu jopa eri lajeiksi. Uudet havainnot osoittavat, ett‰ Daphnia cucullata -vesikirpun "kyp‰r‰n" kokoon vaikuttaa lis‰ksi myˆs vanhempien elinymp‰ristˆ. Er‰‰n Naturessa julkaistun artikkelin [1] mukaan yksilˆn ja sen ‰idin kehitysymp‰ristˆill‰ on suunnilleen yht‰ suuri vaikutus. Kun sek‰ aiti ett‰ yksilˆ itse eliv‰t kairomoniymp‰ristˆss‰, kehittyi kyp‰r‰ l‰hes kaksi kertaa niin suureksi kuin vertailuryhm‰ll‰. Kummankin tekij‰n vaikutus yksin‰‰n aiheutti muutoksen, jonka suuruus oli noin puolet yhteisvaikutuksen aiheuttamasta muutoksesta. Artikkelissa tarkasteltiin myˆs vastaavanlaista muutosta, jonka naurisperhosen (Pieris napi) toukat aiheuttivat peltoretikan (Raphanus raphanistrum) puolustuskemikaalien tuotannossa.
T‰llaisia yhden kerran periytyvi‰ muutoksia tunnetaan ainakin 60 [15]. Muutokset n‰kyv‰t yleens‰ puolustuskyvyss‰ ne aiheuttanutta ymp‰ristˆtekij‰‰ vastaan tai ominaisuuksissa, joihin kyseinen ymp‰ristˆtekij‰ luontaisesti vaikuttaa. (T‰llainen on esimerkiksi tavallista nopeampi kasvu, joka aiheutuu ravinteiden lis‰‰ntymisest‰.) Yhden kerran periytyvi‰ muutoksia on havaittu lukuisissa eri el‰inryhmiss‰ kuten kasveissa, hyˆnteisiss‰, matelijoissa, sammakkoel‰imiss‰ ja jopa nis‰kk‰iss‰ [15]. Nis‰kk‰iden kohdalla t‰ytyy kuitenkin muistaa, ett‰ kemiallisia ominaisuuksia, kuten vasta-aineita ja viruksia, siirtyy runsaasti j‰lkel‰isiin istukan ja ‰idinmaidon v‰lityksell‰ [2].
Yritykset selitt‰‰ dauermodifikaatiota pelk‰st‰‰n geenimutaatioiden avulla kaatuvat ilmiˆn portaattomuuteen ñ muutokset usein syntyv‰t ja h‰vi‰v‰t v‰hitellen. Lis‰ksi olisi varsin ep‰todenn‰kˆist‰, ett‰ perim‰ss‰ tapahtuisi takaisinmutaatioita aina kun muutoksen aiheuttanut ymp‰ristˆtekij‰ poistetaan [9]. Voidaan siis pit‰‰ varsin todenn‰kˆisen‰, ett‰ dauermodifikaatio aiheutuu muutoksesta solulimassa eik‰ geenimutaatioista. Solulimassa tapahtuvaa periytymist‰ onkin tutkittu paljon ñ myˆs sellaisten ominaisuuksien kohdalla, jotka eiv‰t suoranaisesti ole ymp‰ristˆn aiheuttamia [2, 12, 17]. Varhaisten tutkimusten luotettavuutta on kuitenkin vaikea arvioida, sill‰ tuolloin mitokondrio-RNA:n eik‰ plasmidien eli solulimassa vapaina olevien DNA- renkaiden olemassaoloa ei viel‰ tunnettu.
Muutoksen asteen voisi ajatella riippuvan jonkin tietyn kemikaalin tai partikkelin m‰‰r‰st‰ solulimassa. On ehdotettu, ett‰ t‰llainen aine voisi siirty‰ munasolun v‰lityksell‰ j‰lkel‰isiin, jolloin sen vaikutus jatkuisi, kunnes se on solun jakautumisen seurauksena laimentunut tarpeeksi. T‰llainen vaikutus ei kuitenkaan voisi kest‰‰ kovin pitk‰‰n, sill‰ jo 30 solunjakautumisen aikana kemikaali laimenisi miljardisosaan alkuper‰isest‰. Ainakin jotkin vain yhden kerran periytyvist‰ muutoksista voitaisiin kuitenkin mahdollisesti selitt‰‰ t‰ll‰ tavoin. Onkin ehdotettu, ett‰ niiss‰ olisi kysymys aivan eri ilmiˆst‰ kuin Jollosin kokeissa, eik‰ niit‰ siksi pit‰isi lainkaan kutsua dauermodifikaatioiksi [20]. Jos ominaisuuden aiheuttaneet solulimapartikkelit pystyisiv‰t kuitenkin itsen‰isesti kopioimaan itse‰‰n kuten virukset, voisi ominaisuus periyty‰ solulimassa loputtomasti. T‰ll‰ menetelm‰ll‰ toimii todenn‰kˆisesti ainakin tohveliel‰imell‰ havaittu plasmaattinen letaalitekij‰, jota merkit‰‰n kreikkalaisella kirjaimella k (kappa) [20]. T‰m‰n letaalitekij‰n siirtyminen konjugaatiossa el‰imelt‰ toiselle riippuu konjugaation kestosta eli siit‰, paljonko solulimaa sen aikana on ehtinyt siirty‰. Se toiminta ei kuitenkaan tiett‰v‰sti riipu elinymp‰ristˆst‰.
Dauermodifikaation mekanismia etsitt‰ess‰ on huomattava, ett‰ vastaava ilmiˆ tapahtuu yksilˆn kehityksess‰ kudosten erilaistuessa. T‰llˆin sit‰ kutsutaan determinaatioksi. Esimerkiksi ihmisen hermosoluilla ja verisuonen sis‰sein‰m‰n soluilla on t‰sm‰lleen sama perim‰. Kuitenkin hermosolut tuottavat jakautuessaan uusia hermosoluja ja verisuonen sis‰sein‰m‰solut uusia sis‰sein‰m‰soluja. Vaikka hermosolu siirrett‰isiin verisuonisolujen keskelle, se tuottaisi edelleen vain uusia hermosoluja.
Alun perin se, millaiseksi jokin solukko kehittyy, riippuu ymp‰rˆivien solujen vaikutuksesta. Kun jokin alkion solu irrotetaan ja siirret‰‰n toiseen paikkaan, se kehittyy uuden ymp‰ristˆns‰ mukaisesti. T‰t‰ ilmiˆt‰ kutsutaan induktioksi. El‰insoluissa tapahtuu kuitenkin jossakin vaiheessa niin kutsuttu determinaatio. T‰llˆin solujen erilaistumista ohjaava tekij‰ alkaa s‰‰dell‰ itse itse‰‰n, eik‰ kehitys en‰‰ riipu ymp‰rˆivist‰ soluista. Jos determinoitunut jalkasolu siirret‰‰n k‰den paikalle, se kehittyy joka tapauksessa jalaksi [19]. Toisin kuin el‰insoluissa, kasvisoluissa determinaatiota ei yleens‰ tapahdu, vaan solut ovat joitakin poikkeuksia lukuun ottamatta totipotentteja eli kykenevi‰ sopivissa olosuhteissa kasvamaan kokonaiseksi kasviksi.
Geenitoiminnan itses‰‰tely toimii niin kutsutun takaisinkytkenn‰n avulla. Geenin tuottama proteiini tai jokin sen reaktiotuote sitoutuu ep‰aktiiviseen aktivaattoriproteiiniin, joka muuttaa muotoaan ja voi sen j‰lkeen toimia helpottaen RNA-polymeraasin kiinnittymist‰ geenin operaattorialueelle. Vaihtoehtoisesti se voi inaktoivoida repressori- eli est‰j‰proteiinin, joka muutoin sitoutuisi geenin promoottorialueeseen est‰en geenin transkription eli tulkinnan. Kummassakin tapauksessa kemikaalia syntetisoidaan jos, ja vain jos, sit‰ jo ennest‰‰n on solussa l‰sn‰. M‰‰r‰n kasvaminen liian suureksi estyy vastaavan, geenitoimintaa hillitsev‰n prosessin avulla.
Saksalainen biologi Alfred K¸hn esitt‰‰ perinnˆllisyystieteen oppikirjassaan [13], ett‰ dauermodifikaatio voisi selitty‰ ns. predeterminaation eli ennen munasolun hedelmˆittymist‰ tapahtuvan determinaation avulla. H‰nen mukaansa predeterminaatio voi aiheutua joko ymp‰ristˆn suorasta vaikutuksesta, ‰idin elimistˆss‰ olevista kemikaaleista tai munasolun omasta perim‰st‰ ñ ennen meioosiahan munasolussa on viel‰ ‰idin koko perim‰ tallella. N‰ist‰ ensimm‰inen voisi selitt‰‰ yhden kerran periytyv‰t muutokset ja toinen myˆs pitemm‰t dauermodifikaatiot, kuten Habrobracon juglandiksella havaittu.
K¸hn ei kuitenkaan ota huomioon sit‰ mahdollisuutta, ett‰ munasolu voisi olla jo valmiiksi determinoitunut. Jokin koko organismiin vaikuttava ymp‰ristˆtekij‰, kuten l‰mpˆtila tai kemikaalit, voisi aiheuttaa determinaation yksilˆn kaikissa, myˆs iturataan kuuluvissa soluissa. Ituradassa tapahtunut determinaatio siirtyisi sitten yksilˆpolvelta toiselle samoin kuin somaattisten solujen determinaatio siirtyy solusukupolvelta toiselle.
T‰st‰ tuskin kuitenkaan on kysymys monisoluisten eliˆiden dauermodifikaatioissa, sill‰ ne kest‰v‰t yleens‰ vain muutaman sukupolven, ja muutoksen v‰hitt‰inen poistuminen edellytt‰isi paljon monimutkaisempaa s‰‰telyj‰rjestelm‰‰ kuin edell‰ kuvattu. Yksisoluisilla sukupolvien v‰linen determinaatio on kuitenkin potentiaalinen ehdokas dauermodifikaatiomekanismiksi. Niill‰ yksilˆn jakautuminen voidaan nimitt‰in suoraan rinnastaa monisoluisilla tapahtuvaan somaattisten solujen jakautumiseen. Itse asiassa tohveliel‰inkloonia voidaan jopa pit‰‰ yhten‰ el‰imen‰, jolloin konjugaatiossa s‰ilyv‰ dauermodifikaatio voitaisiin rinnastaa monisoluisten dauermodifikaatioon.
Yksisoluisillakin muutos h‰vi‰‰ v‰hitellen, joskin pitk‰n ajan kuluessa. Vaikka yleens‰ uskotaankin, ettei dauermodifikaation h‰vi‰minen johdu normaalin ymp‰ristˆn kumoavasta vaikutuksesta, olisi mielest‰ni varsin mahdollista, ett‰ esimerkiksi tohveliel‰imille olisi kehittynyt kyky sopeutua molempiin suuntiin.
Vaikka tietyt el‰m‰n aikana hankitut ominaisuudet voivatkin siirty‰ j‰lkipolville, dauermodifikaatiossa ei kuitenkaan ole kysymys hankittujen ominaisuuksien periytymisest‰ siin‰ mieless‰ kuin Lamarck sen ajatteli. Dauermodifikaatiot eiv‰t voi osallistua lajin kehitykseen, sill‰ ne h‰vi‰v‰t aina ennemmin tai myˆhemmin. Lis‰ksi dauermodifikaatio ilmenee vain tietyiss‰ ominaisuuksissa ñ sellaisissa, joiden s‰‰telyj‰rjestelm‰‰n se on erikseen evolvoitunut. Monissa tapauksissa dauermodifikaatio on selv‰stikin luonnon valinnan avulla saavutettu sopeutuma jatkuvasti muuttuvaan elinymp‰ristˆˆn. Esimerkiksi tohveliel‰inten kyky sopeutua erilaisiin l‰mpˆtiloihin ilman geeniperim‰n muutosta varmasti parantaa lajin elinkelpoisuutta. Samoin vesikirpulle on hyˆty‰ siit‰, ett‰ se jo el‰m‰ns‰ alussa tiet‰‰ mahdollisesta vaarasta ja voi aloittaa valmistautumisen heti. Tarhapavun dauermodifikaatiota on kuitenkin vaikeampaa selitt‰‰. Kuinka kasvuh‰iriˆiden periytyminen voisi olla lajille edullista?
Solulimaperiytyminen kuitenkin mit‰ ilmeisimmin osallistuu evoluutioon, vaikka sen osuus ei liene l‰hesk‰‰n yht‰ merkitt‰v‰ kuin geenien. Solulima sis‰lt‰‰ paljon geenien toiminnalle v‰ltt‰m‰tˆnt‰ informaatiota. Muutokset epigeneettisess‰ eli geenien transkriptiosta eli tulkinnasta vastaavassa systeemiss‰ luonnollisesti vaikuttavat geenien toimintaan, eik‰ edes standardina pidetty geneettinen koodi eli DNA:n em‰sten ja aminohappojen v‰linen vastaavuus ole kaikilla eliˆill‰ sama. Joidenkin tutkijoiden mukaan epigeneettisess‰ systeemiss‰ tapahtuvilla muutoksilla on paljon luultua suurempi osuus evoluutiossa [5]. Soluliman merkityst‰ lajien v‰lisiss‰ eroavaisuuksissa on vaikea testata, sill‰ eri lajien yksilˆt eiv‰t yleens‰ risteydy kesken‰‰n. On kuitenkin selv‰‰, ett‰ vaikka tohveliel‰imeen siirrett‰isiin ihmisen perim‰, se ei kehittyisi ihmiseksi.
Vaikka sukupolvien v‰linen determinaatio ei selv‰stik‰‰n aiheuta monisoluisten eliˆiden dauermodifikaatiota, se saattaa kuitenkin selitt‰‰ varsin suuren osan solulimaperiytymisest‰. Jos otetaan huomioon, kuinka yleinen ilmiˆ determinaatio on kudosten erilaistumisessa, voisi kuvitella, ett‰ kaikilla eliˆill‰ on lukuisia ominaisuuksia, jotka ovat s‰ilyneet soluissa îdeterminoituneinaî ñ geneettisen takaisinkytkenn‰n avulla ñ jo miljoonien vuosien ajan ja s‰ilyv‰t edelleen niin kauan kuin se on lajille edullista. N‰m‰kin ominaisuudet samoin kuin dauermodifikaatiokyky sek‰ muutokset epigeneettisess‰ systeemiss‰ syntyv‰t mit‰ ilmeisimmin luonnonvalinnan eiv‰tk‰ siis lamarckismin mukaisesti ymp‰ristˆn suuntaavan vaikutuksen tuloksena.
Jostakin syyst‰ modernit kehitysbiologian ja perinnˆllisyystieteen oppikirjat ja hakuteokset eiv‰t mainitse dauermodifikaatiota lainkaan. Tiedossani ei ole yht‰‰n mainintaa varsinaisesta dauermodifikaatiosta tieteellisess‰ kirjallisuudessa vuoden 1971 j‰lkeen [13, 14]. Yhden kerran periytyvist‰ muutoksista on tosin myˆhemminkin tehty lukuisia tutkimuksia maternaaliefektin nimell‰ [15].
Referoimani tutkimukset ovat kuitenkin mielest‰ni olleet perusteellisia, eik‰ ilmiˆn olemassaoloa ole syyt‰ ep‰ill‰. En ole myˆsk‰‰n lˆyt‰nyt yht‰‰n vakavasti otettavaa kritiikki‰. Vaikka dauermodifikaatio onkin mit‰ ilmeisimmin todellinen ilmiˆ, siin‰ ei kuitenkaan miss‰‰n tapauksessa ole kysymys lamarckismista, kuten er‰‰t kirjoittajat (esim. [4]) tuntuvat luulevan. Dauermodifikaatio ei myˆsk‰‰n ole mill‰‰n tavoin ristiriidassa evoluutioteorian kanssa, sill‰ muutokset ovat aina geenitoiminnan s‰‰telemi‰. Dauermodifikaatiosta ei kuitenkaan tiedet‰ paljoa. On s‰‰li, ettei siit‰ ole tehty enemp‰‰ tutkimusta.
| ||||||||||||||||||||||||||||||
|
Taulukko 1: Muutamia merkitt‰vimpi‰ esimerkkej‰
dauermodifikaatiosta. |
[1] Agrawal, Anurag A.; Laforsch, Christian & Tollrian, Ralph; Transgenerational induction of defences in animals and plants. Nature 401 (1999): 60-63.
[2] Caspari, Ernst; Cytoplasmatic inheritance. Advances in Genetics 2 (1948).
[3] Caspari, Ernst; The role of genes and cytoplasmic particles in differentiation. Annals of the New York Academy of Sciences 60 (1955).
[4] Custance, Arthur C.; Science and Faith. The Doorway Papers Series 1988. http://www.custance.org/Library/Volume8/index.html
[5] Ho, M.W. & Saunders, P.T.; Beyond neo-darwinism ñ An epigenetic approach to evolution. Journal of Theoretical Biology, 78 (1979): 573-591.
[6] Hofmann, Frederick Wenzl; Some attempts to modify the germ plasm of Phaseolus vulgaris. Genetics 12 (1927): 284- 294.
[7] Jollos, Victor; Experimentelle Untersuchungen an Infusorien. Biologisch Zentralblatt. 33 (1913): 222-236.
[8] Jollos, Victor; Experimentelle Protistenstudien ñ I. Untersuchungen ¸ber Variabilit‰t und Vererbung bei Infusorien. Archiv f¸r Protistenkunde 43 (1921): 1-222.
[9] Jollos, Victor; Dauermodifikationen und Mutationen bei Protozoen. Archiv f¸r Protistenkunde 83 (1934): 197- 219.
[10] Jollos, Victor; Inherited changes produced by heat-treatment in Drosophila melanogaster. Genetica 16 (1934): 476- 494.
[11] Kaestner, Hans; Die Wirkung von Temperaturreizen auf die Pigmentierung und Ihre Nachwirkung in den folgenden Generationen bei Habrobracon juglandis Ash. Wilhelm Roux' Archiv f¸r Entwicklungsmechanik der Organismen 124 (1931).
[12] King, Karl; Cytoplasm, Inheritance and Mutations. 2001.
[13] K¸hn, Alfred; Grundriþ der Vererbungslehre. Quelle & Meyer, Heidelberg 1971.
[14] K¸hn, Alfred; Lectures in Developmental Physiology. Springer-Verlag, 1971.
[15] Rossiter, MaryCarol; Incidence and consequences of inherited environmental effects. Annual Review of Ecology and Systematics 27 (1996): 451-476.
[16] Schuckmann, W. von & Piekarski, G.: Beitr‰ge zum Problem der Dauermodifikation bei Protozoen. Archiv f¸r Protistenkunde 93 (1940): 355-416.
[17] Sirks, M. J.; Plasmatic inheritance. The Botanical Review IV/3 (1938): 113-131.
[18] Schlottke, E.; Ðber die Variabilit‰t der schwarzen Pigmentierung und ihreBeeinfluþbarkeit durch Temperaturen bei Habrobracon juglandis. Zeitschrift f¸r Vergleichende Physiologie 3 (1926).
[19] Walbot, Virginia; Holder, Nigel; Developmental Biology. Random House, 1987.
[20] Wichterman, Ralph; The Biology of Paramecium. New York, 1953.